Musculature and Exoskeleton
The hard, protective exoskeleton (or cuticle) is perhaps the most immediately obvious feature significantly separating ourselves and other vertebrates from arthropods such as hermit crabs. The exoskeleton is composed of non-living material, much like the hair or nails of mammals, and is secreted by the epidermis (Ruppert et al. 2004). The epidermis, the ‘skin’ of mammals, is derived from the outer most of the three primary tissue layers of the embryo, the ectoderm, and in arthropods lies directly beneath the exoskeleton (Ruppert et al. 2004).
|
|
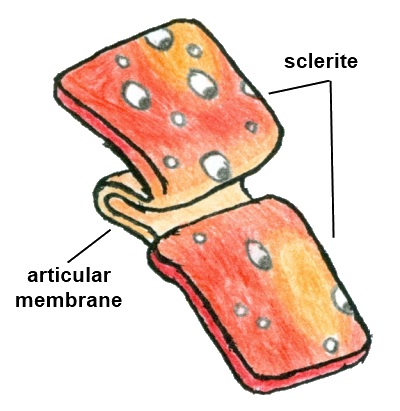
Stylisation of hardened, protective sclerite plates with soft, flexible articular membrane hidden below. Illustration: Storm Martin 2012, adapted with modification from Ruppert et al. 2004
|
The cuticle is composed of chitin, a polysaccharide (carbohydrate), and proteins. Despite most familiarly being hard and rigid, the exoskeleton, when first secreted, is soft (Ruppert et al. 2004). The formation of covalent bonds between the protein components hardens the cuticle, a process known as sclerotisation (Ruppert et al. 2004). After initial hardening by sclerotisation, crustaceans further add to the strength of their cuticle by incorporating inorganic calcium carbonate. This is called biomineralisation, or more precisely described as calcification (Lowenstam and Weiner 1989). Biomineralisation is not a process unique to the crustaceans. Many marine invertebrates incorporate inorganic minerals to produced hardened structures for support and defence, the most notable of which are the hard corals comprising the reef itself and the gastropod molluscs upon which hermit crabs depend (Lowenstam and Weiner 1989). Calcifying organisms such as crustaceans make use of calcium and carbonate ions, both of which are most readily available in more alkaline, warmer waters (Lavali and Spanier 2007). Under these conditions calcium carbonate salts are more insoluble, forming two common crystalline structures; calcite and aragonite (Lavali and Spanier 2007), only the former of which is incorporated into the crustacean cuticle (Lowenstam and Weiner 1989). Crustaceans may also incorporate amorphous calcium carbonate, and malacostracans in particular do so in appreciable quantities (Lowenstam and Weiner 1989).
Some regions of the cuticle do not undergo hardening to the extent of others, remaining soft and flexible (Ruppert et al. 2004). These areas are of course vulnerable to attack from predators and so are not obvious, concealed as folds of the cuticle underneath protective sclerotised regions (Ruppert et al. 2004). Combining flexibility with protectiveness, the exoskeleton is then a series of overlapping sclerotised plates, called sclerites, connected by flexible, unsclerotised cuticle known as the articular membrane (Ruppert et al. 2004). Typically each body segment of an arthropod is framed by four sclerites; the dorsal tergite, the ventral sternite and two lateral pleurites (Ruppert et al. 2004). The carapace of crustaceans is formed by the fusion of several tergites into a continuous protective covering (Ingle and Christiansen 2004). For hermit crabs, the additional protection of a gastropod shell allows for a reduction in sclerotisation and mineralisation of the abdomen (Chockalingam 1974). Indeed, reduced calcification of the abdomen is required in order to permit the coiling into the shell spiral (Ingle and Christiansen 2004). While the abdomen is soft, the cephalic sclerites remain fully calcified, and the thoracic sclerites are only partially hardened (Chockalingam 1974).
A consequence of possessing such well developed armour is the requirement for interaction between the external and internal environments to take place via specialised cuticular structures (Garm 2004). Of the exoskeleton of decapods, such structures are abundant, diverse and may be highly complex in form at the microscopic level. These include spines, simple and sharp projections of cuticle which afford some protection from predators and can be particularly important in the survival of planktonic larvae (Ruppert et al. 2004). Teeth are another class of sharp, rigid projections. In many decapods, including hermit crabs, teeth are most common about the mouthparts and along the biting edge of the chelae (Garm 2004). Hermit crabs use the smaller cheliped to investigate new host shells and the teeth of the chelae have been demonstrated to have a mechanosensory role, aiding in this investigation and at other times associated with food manipulation (Mesce 1993). There also exist a variety of more minute ornamentations including tubercles, pits and pores (Mesce 1993, Garm 2004). Perhaps the most important projections however, are setae, typically defined as articulated chitinous projections filled with the cytoplasm of a single epidermal cell, which spans the cuticle (Garm 2004). In more common terms these are the hairs or bristles, which of D. megistos are very obvious, covering most of the body aside from the smooth abdomen. Setae are highly diverse in structure and microstructure and serve a variety of important physiological roles (Garm 2004). The mouthparts house a particularly diverse and dense assemblage of setae, establishing filter feeding currents, sorting and manipulating food and providing chemosensory and tactile information.
|
|
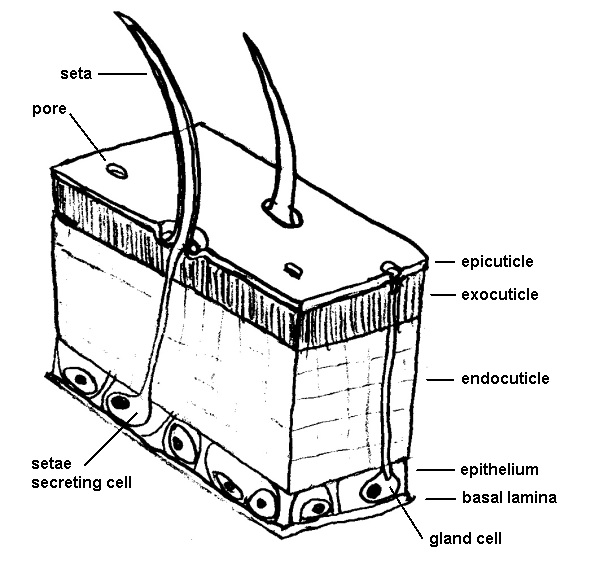
The layers of the cuticle, note relative thickness and trans-cuticle setae secreting cell. Illustration: Storm Martin 2012, adapted with modification from Ruppert et al. 2004
|
The exoskeleton itself is, at a finer scale, composed of three distinct layers, from distal (outer) to proximal (inner): the epicuticle, exocuticle and endocuticle, the latter underlain by the epidermis (Roer and Dillaman 1984, Lowenstam and Weiner 1989, Ruppert et al. 2004). In decapods, the epicuticle is very thin, up to 5µm thick (Roer and Dillaman 1984), is composed of lipoproteins and calcite, but not chitin (Ruppert et al. 2004) and is sparsely mineralised (Lowenstam and Weiner 1989). The exocuticle and endocuticle are both much thicker than the epicuticle, with the endocuticle the thickest of all, two to three times that of the exocuticle (Roer and Dillaman 1984). Both contain chitin and protein, but at different ratios; about a third of the organic material of the exocuticle is chitin, compared with around three quarters in the endocuticle (Roer and Dillaman 1984). The exocuticle is hence the primary site of sclerotisation (Roer and Dillaman 1984), indeed the endocuticle is not sclerotised at all, instead the primary site of calcification (Lowenstam and Weiner 1989).
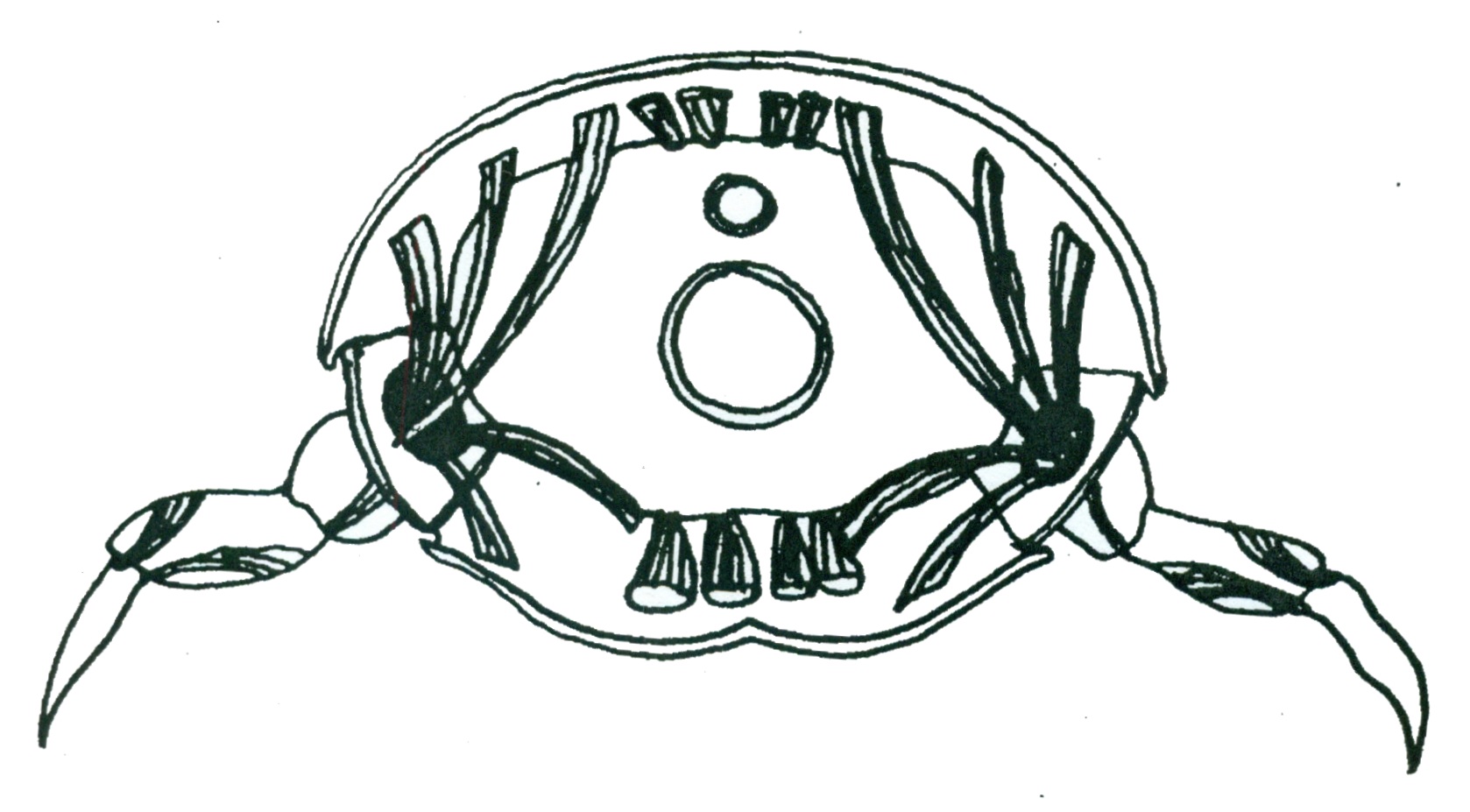
Another consequence of an external skeleton is the necessity of internal muscle attachment. Muscles attach to grooves or ingrowths of the cuticle termed apodemes (Pilgrim 1973). Muscles of the appendages attach through apertures of the pleurites to the inner body wall (Pilgrim 1973). The pereopod apertures of hermit crabs, particularly the first three pairs, are relatively large(Pilgrim 1973).
|
|
Representation of arthropod musculature showing attachment of muscles to internal side of cuticle. Illustration: Storm Martin 2012, adapted with modification from Ruppert et al. 2004
|
The exoskeleton also limits growth, requiring periodic molting of the entire cuticle, including the foregut,
hindgut, infolded articular membranes and appendages. In all arthropods this process is known as ecdysis (Ruppert et al. 2004). During the molt the hermit crab is vulnerable and most susceptible to mortality. Larger individuals progressively molt less and less frequently as they grow. Molting not only allows knew growth but also regenerates lost appendages. Before molting the hermit crab becomes less active and may bury or otherwise hide itself. After molting the hermit crab remains inactive, allowing its new skeleton to harden. |