Ecology
Little is known of the specific ecology of R. graeffei, which is somewhat surprising due to its abundance on Heron Island Reef and presumably throughout the Great Barrier Reef (see FUTURE RESEARCH). There have been no studies on feeding preferences, predators, associations, or distribution patterns. Thus the ecological information presented is based on higher taxonomic levels, most of which comes from studies of encrusting Cheilostomes, particularly in the reefs of the Caribbean. In these Caribbean studies, they found a high degree of heterogeneity in distribution of cheilostomes, with a propensity for a small number of irregularly spaced but massive colonies instead of regularly spaced smaller colonies (Jackson 1984). This makes sense in the context of a different study by Winston and Jackson (1984) in which they showed recruitment and growth of Cheilostome colonies to be far slower in their tropical study site than reported in temperate waters. Thus they found that size of a colony was directly related to maximum survival. Both the distribution and physical structure of the colonies were also influenced by grazing patterns of damselfish.
In addition to pomacentrid fishes, bryozoans are known to be predated upon by nudibranchs, which are able to drill through the calcareous zooecium with their radula and use their pharyngeal pump to extract the zooid (Miller 1961). However, colonies are able to respond to chemical cues in their environment fairly rapidly (within days) and studies of Membranipora membranacea showed a remarkable ability to grow defensive corner and membranous (from the frontal membrane) spines when presented with chemical cues from different nudibranch species (Iyengar & Harvell 2002).
Pycnogonids are often found on bryozoan colonies, and appeared on specimens of R. graeffei collected from Heron Island.
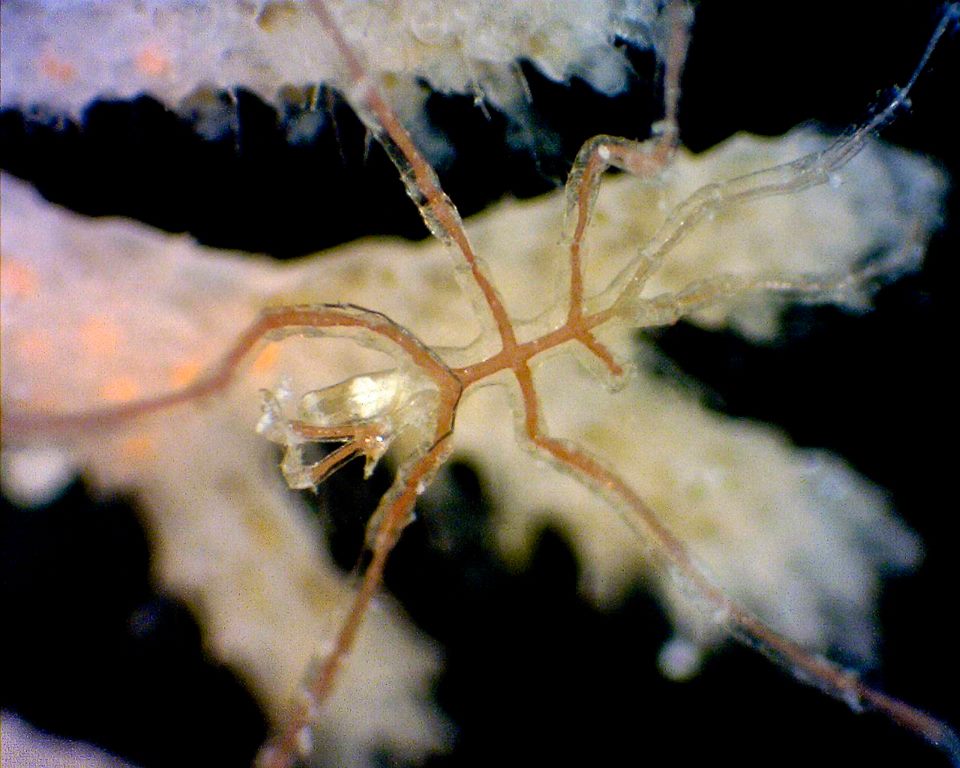
|
|
Pycnogonid sp. found on R. graeffei spciemen. Photo: Bridget Bradshaw, Heron Island Reef, 2013. |
However, their relationship with the bryozoans is not well known. An Antarctic species of pycnogonid (Austrodecus glaciale) is thought to use its long proboscis to probe inside the zooecium through the pseudopores of ascophorans, though the act of feeding has never been directly observed (Fry 1965). Urchins, such as Diadema antillarum, have been observed grazing under corals and causing lesions in the bryozoan colonies living there (Jackson & Winston 1981). While bryozoans are able to quickly bud and fill these lesions with new zooids, there is strong competition for newly freed space with sponges and hydroids.
In terms of feeding ecology,bryozoans all use an upstream suspension-feeding collecting system and do not require light to grow, but rather nutrient rich water. Most feed on phytoplankton or the occasional bacteria or detritus that falls into their ring of tentacles (Ryland 1970).
|